Formation of stable patterns by local autocatalysis and depletion of a long-ranging substrate
An alternative to the activator - inhibitor mechanism described above is the antagonistic effect results from a depletion of a substrate or co-factor that is required for the self-enhancing reaction (Gierer and Meinhardt, 1972 ). Most inorganic pattern-forming systems such as the formation of sand dunes and patterns of erosion depend on such a depletion mechanism. Assumed in the simulations below is a substance that feeds back to increase its own concentration (activator, green). This local autocatalysis is balanced by the depletion of a substrate (red) required for the autocatalytic reaction. In a region that has an extension comparable with the range of the activator (i.e., in fields much smaller than the range of the substrate), only a single maximum is generated at one marginal position. Minute fluctuations are sufficient to initiate patterning formation After removal of the activated region, the substrate concentration rises steeply in the remaining field, causing the polar pattern to regenerate:


n a somewhat larger field, only symmetrical patterns are possible with either a maximum at each end or a single maximum in the center. The left figure shows the initial condition, two intermediate and the final pattern (equations and parameters ):

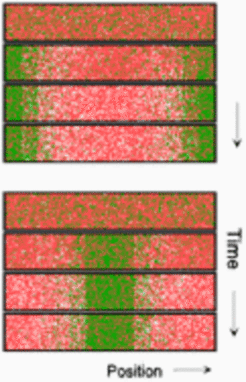

In small but growing fields, a marginal maximum forms first. Upon further growth, the substrate concentration in the non-activated part increases. The substrate concentration at the flank of the maximum can become so high that the autocatalysis proceeds there more rapidly than in the center of the maximum. The position of the peak becomes instable and shifts to the center. For the same reason, a maximum splits into two, and so on, during further growth:


In contrast, if the antagonistic reaction results from an inhibitor that is produced under activator control, periodic patterns form by insertion of new maxima into the enlarging interstices, rather than by splitting of existing maxima (see Insertion of new maxima ; for more details of this behavior, see Meinhardt, 2003 ). The different behavior is based on an upper bound of the self-enhancing reaction inherent in the activator - depleted substrate system since the activation can no longer increase if all precursor molecules are used up. After introduction of a saturation in an activator-inhibitor system, both systems behave very similar. A feature of satuating systems that is important for many biological applications is that maxima can easier move to a preferential position and they can split.

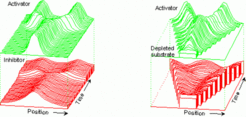
Such a peak splitting is well known in the generation of branching structures during ontogenetic development. For instance, during lung development maxima of the growth factor FGF direct the outgrowth of branches. If sufficient space is available, such peaks can split. According to the theory the reason is as follows: if the self-enhancement saturates at higher levels of the self-enhancing reaction the maximum peak height is restricted and a broadening occurs instead. Cell at the flanks of a peak can be in a better position than those in the centre since in the centre not only the self-enhancement but also the antagonistic effects are highest. Thus, broadening of a peak, e.g., by a minor increase in the saturation can lead to a higher 'reproduction rate' of the auto regulatory molecules at the flanks than at the centre, causing a peak to split (left figure). Likewise, peak splitting can occur in an activator-depleted substrate interaction if more resources become available, e.g., due to growth (right figure and simulations above). Such a peak splitting is proposed to be the reason if two midlines are formed in a Drosophila mutant.