Activation of genes under the influence of morphogenetic signalling
Autoregulatory feedback loops are required
The generation of signals by the exchange of molecules via diffusion works only in small fields. In larger fields the time required to generate patterns by randomly moving molecules would be much too long. Therefore, the signals generated at small scales have to be translated into more permanent states of differentiation that can be maintained upon further growth. The obvious means is a stable concentration- (and thus space-) dependent activation of genes. The choice of a particular pathway under the influence of a morphogenetic signal requires the activation of particular genes and the suppression of alternative genes. This situation has formal similarities with pattern formation in space: spatial patterning requires activation at a particular position and the inhibition in the remaining part. In analogy, the selection of a particular pathway requires the activation of a particular gene and the suppression of the alternative genes. Based on this similarity, I have proposed in 1978 that gene activation requires a direct or indirect feedback of genes on their own activation and their mutual competition such that only one of the alternative genes can remain active in a particular cell. [ PDF ]. Meanwhile many such autoregulatory genes have been found. The genes deformed [8], hunchback or twist [10] are examples. As in pattern formation, the feedback has to be non-linear . This condition is satisfied if dimers are involved in the autoregulation.

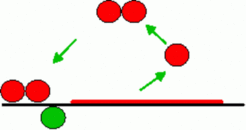

Genes with an a positive autoregulatory feedback can acts as a switch. Whenever the signal is above a threshold, the gene activity switches from the OFF- into the ON-state. Due to the autoregulation, the response of the cell becomes in a wide range independent of the exact morphogen concentration. Therefore, small deviation from a desired signaling strength are not propagated into the next hierarchical level of gene activation. The signal is required only for the initiation, not for the maintenance of gene activity, i.e., the cells obtain some sort of a long-term memory in respect to the signals they have seen ( for equation click here ).
Unidirectional promotion: activation of several genes by a single gradient
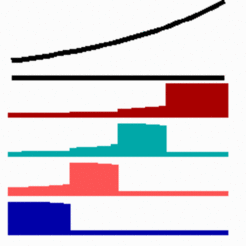
For the position-dependent activation of of several genes by a single gradient genes I have proposed that the cells do not measure a particular morphogen concentration all at ones. Instead, starting from a default gene activity, other genes become activated in a unidirectional manner. Each further step requires a higher morphogen concentration. This process comes to rest if the actual gene activation corresponds to the local morphogen concentration.

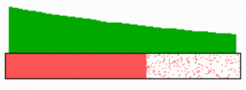
The simulation above shows the activation of four genes by a morphogen gradient (black, top). Starting with a default gene (blue), a 'promotion' from one gene to the next leads to sharply confined regions in which particular genes are active. Such a model predict that the first transition from one gene activation to the next occurs in the region of the highest morphogen concentration. As can be seen with the 'red' gene in this simulation, the activation sweeps over the field until it comes to rest in a defined region (for equation click here).
The model is based on the following assumptions
- The gene products of the genes to be activated by the graded signal have a (direct or indirect) positive non-linear feedback on the activation of their own gene.
- The genes mutually repress each other such that only one gene of the set can be active in a particular cell.
- Cells do not measure different levels in a single step but they compare their achieved state of determination with the strength of the external signal. A sequential transition from one gene activation to the next occurs as long as the signal is still high enough. Each of the subsequent activations requires a higher concentration and each step requires a certain time.
Note that in this mode of gene activation the cells compare only their state of determination with the external signal. The cells do not communicate with each other whether they are the correct neighbors. Therefore, a later induced perturbation in the neighborhood can neither be detected or repaired. This is in strong contrast to patterning within segments.
An analogy
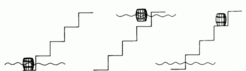
The situation can be compared with a barrel at the base of a staircase. The barrel may be lifted up by a flood (morphogen signal). After lowering of the flood, the barrel can only remain at a few discrete levels (activation of particular genes). A later, higher flood can lift the barrel up even further; a second lower flood would have no effect.

A premature removal of the morphogen leads to an arrest in the promotion ( simulation below). Anterior structures remain at a more posterior position while the most posterior structures could be missing. The specification of digits in the chicken wing under the influence of the zone of polarizing activity (ZPA) has this dynamics.


Meanwhile several systems with unidirectional promotion have been observed. A low concentration of Activin (a member of a family of growth factors) causes in animal cup cells of Xenopus the activation of the Xbra gene while for the gene Xgsc a higher concentration is required. Applying Activin first in low and later in high concentrations leads to a reprogramming of cells from Xbra to Xgsc, while after a later reduction is without effect [1].
This mode of unidirectional promotion has a very essential function during development. As mentioned, the range of the signalling molecules is usually small. The initial activation of genes takes place in small fields that grow. During this growth, the distance between a cell and a morphogen source increases. Thus, the morphogen concentration necessarily decreases. Due to this unidirectional character of the promotion, the cell maintain their once obtained state of differentiation. This stability is possible without that the determination is absolutely fixed. If, for instance, during regeneration, a new morphogen source is generated, the cells can still be 'distally transformed'. This plays a decisive role, for instance, in the regeneration of cockroach legs The unidirectional promotion enables on the one hand the required stability to cope with the fading influence of the morphogen during growth and provides, on the other, still some (unidirectional) flexibility if regulation is required.
The model predicts that genes that become activated in regions of low morphogen concentration are more sensitive to this signal. The question is then how it can be achieved that the less sensitive genes have nevertheless their chance in an region of high morphogen concentration. The expectation of from the theory is that the 'higher' genes have a more efficient autoregulation such that they win at high concentration in spite of the lower sensitivity.
Why antagonists of morphogens could be essential
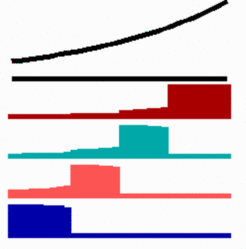
In Xenopus there is good evidence that the pattern of the head is set up by a Wnt signal that spreads from the marginal zone [4,5]. Low Wnt concentrations are necessary for the formation of the most anterior structures. Most remarkably, in this region many WNT antagonists are present [6,7], partially supplied under maternal control. If missing, anterior structures (forebrain) will not form. Why is a signal produced that quench the function of other molecules, at least for a substantial period? The model outlined above provides a rational. If a gradient is generated by an pattern-forming system as described above, it may take some times until the concentration opposite to the organizing region has obtained its low steady state level. The same is true if the field is originally so small that the gradient level at the side antipodal to the organizer is too high to determine the most anterior structures. These structures will be missing (blue and magenta gene in the simulation below):

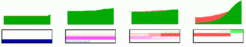
However, if an antagonist quenches the signal for posterior transformation for a certain period, the low concentration can be achieved before the irreversible promotion starts. In the simulation below, the (Wnt)-antagonist (blue bar) blocks the promoting action of the morphogen (activator, green). After its degradation the promotion starts and also the "most anterior genes" (blue and magenta) become expressed:


The proposed mode of gene regulation predicts that cells adapt to the new environment after transplantation into a region of higher morphogen concentration. In the simulation below after transplantation of a cell with the 'low ' gene 1 (blue) active into a region of high signal concentration, gene 4 (brown) becomes becomes activated.


In contrast, cells transplanted from a region of high into a region of low morphogen concentration maintain their differentiation:

A corresponding behavior has been observed after transplantation of cells of the hindbrain [3,4]. Retinoic acid mimics the natural morphogen. A transplantation to a posterior position, i.e., towards the presumptive morphogen source, leads to an adaptation to the new position, while a the pattern of gene activation is maintained after a transplantation to a more anterior position, i.e. away from the source. Likewise, transplantation of neuronal precursor cells of the central nervous system in Drosophila adapt after transplantation towards the ventral midline but remain but appear finally determined after transplantation in the opposite direction.
Further Reading and References
Original paper: Meinhardt, H. (1978). Space-dependent cell determination under the control of a morphogen gradient. J. theor. Biol. 74, 307-321 [PDF]
- Gurdon, J.B., Mitchell, A. and Mahony, D. (1995). Direct and continuous assessment by cells of their position in a morphogen gradient. Nature 376, 520-521.[2]Gould, A., Itasaki, N. and Krumlauf, R. (1998). Initiation of rhombomeric HoxB4 expression requires induction by somites and a retinoic acid pathway. Neuron 21, 39-51.
- Grapin-Botton, A., Bonnin, M.A., Sieweke, M. and Le Douarin, N.M. (1998). Defined concentrations of a posteriorizing signal are critical for MadB/Kreisler segmental expression in the hindbrain. Development 125, 1173-1181.
- Kiecker, C. & Niehrs, C. (2001). A morphogen gradient of wnt/beta-catenin signalling regulates anteroposterior neural patterning in Xenopus. Development 128, 4189-4201.
- Dorsky, R.I., Itoh, M., Moon, R.T. & Chitnis, A. (2003). Two tcf3 genes cooperate to pattern the zebrafish brain. Development 130, 1937-1947.
- Glinka, A., Wu, W., Delius, H., Monaghan, A.P., Blumenstock, C. and Niehrs, C. (1998). Dickkopf-1 is a member of a new family of secreted proteins and functions in head induction. Nature 391, 357-362.
- Bouwmeester, T., Kim, S.H., Sasai, Y., Lu, B. and De Robertis, E.M. (1996). Cerberus is a head-inducing secreted factor expressed in the anterior endoderm of Spemanns organizer. Nature 382, 595-601.
- Regulski, M., Dessain, S., McGinnis, N. and McGinnis, W. (1991). High-affinity binding-sites for the deformed protein are required for the function of an autoregulatory enhancer of the deformed gene. Genes Dev. 5, 278-286
- Simpson-Brose, M., Treisman, J. and Desplan, C. (1994). Synergy between the hunchback and bicoid morphogens is required for anterior patterning in Drosophila. Cell 78, 855-865.
- Leptin, M. (1991). twist and snail as positive and negative regulators during Drosophila mesoderm development. Genes Dev. 5, 1568-1576.