Left-right pattern: the midline squeeze a second pattern to the side
Considerable progress has been made in the understanding of the left-right pattern (for review see [1,2,8]). In the following, it will be shown that many observation can be accounted for by the assuming that the midline system induces at long range and represses at short range a second system that marks the left side. Therefore, the ‘left‘ system depends on the midline system, but is shifted to the side. The following simulation shows the pattern in a cross-section perpendicular to the midline.




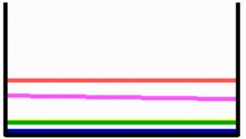
First a high concentration of an activator emerges that marks the midline (green, see simulation on the preceding page). It is formed at the position where the competence for this is highest (dotted curve). This induces on longer range a second patterning system that marks 'left' (blue). Since the latter is locally suppressed by the midline system, it is squeezed to both sides. Due to the long-ranging competition, a full 'left'-maximum can develop only at one side. A minute asymmetry (solid curve) is sufficient to determine which side will win. After removal of the left side, first the midline marker regenerates. This triggers again the 'left' signal (blue). It is shifted now to the right since the left side no longer exists. This simulation is in agreement with the observation [3]. A movie shows the concentration profiles as curves.
The left-right patterning is an obvious feature of vertebrate development but not in insects [1]. The different mechanisms discussed above for midline formation provides a rational why a left-right pattern is easier to realize in vertebrates. Since midline elongation occurs in vertebrates by the spot-like node / organizer, a displacement of the left signal can lead to an unambiguous decision. In the following simulation for the L-R signal in vertebrate, the organizer (green) elongates the midline (red). Midline and organizer induce the 'left'- signal realized by the Nodal/Lefty2 (blue) system that becomes excluded from the midline. Due to the long-ranging inhibition, the 'left'- signal can only appear on one side of the node.

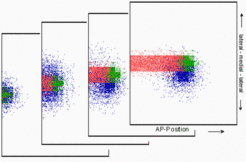
In contrast, since the insect midline has from the beginning a long AP extension, a squeezing out of the 'left'-signal by the midline would not lead to a signal that is consistently located on one site. In some regions it can appear on the left and in the others on the right side. Such a mosaic character would be enforced by the lateral inhibition mechanism inherent in the Nodal/Lefty2 system (shown are subsequent stages).


In Siamese twins fused side-by-side, one of the embryos has frequently a situs inversus. In the animated simulation below the two "left" signals (blue) appear on different sides of the two midline signals (green) although both are exposed to the same bias (oblique pink line). In other words, the mutual inhibition is stronger than the bias.

The model is compatible with several observations. Nodal, the key gene for the 'left' signal, has a positive feedback on its own activation [4,5]. It also regulates the production of a putative inhibitor, lefty-2 [6]. The repression of nodal by the midline system, i.e., the notochord, has been shown (Lohr et al., 1998). Many observations demonstrate an important role of the midline signal in the generation of the left-right pattern [7]. Excised tissue from the right side that does not contain notochord and, of course, no activity characteristic of the left side, regenerates first notochordal maker and later nodal, the marker for the left side [3]. Thus, many of the essential ingredients required by the model seem to be present.
Further Readings and References
Meinhardt H. (2001). Organizer and axes formation as a self-organizing process. Int. J. Dev. Biol. 45:177-188 [PDF]
- Cooke J. (2004). The evolutionary origins and significance of vertebrate left-right organisation. BioEssays 26: 413-421.
- Rasskin-Gutman D, Izpisúa-Belmonte JC.(2004). Theoretical morphology of developmental asymmetries. BioEssays 26: 405-412.
- Levin, M. and Mercola, M. (1998). Evolutionary conservation of mechanisms upstream of asymmetric nodal expression: reconciling chick and Xenopus. Dev. Genetics 23, 185-193.
- Schier, A.F. and Shen, M.M. (2000). Nodal signaling in vertebrate development. Nature 4, 385-389.
- Saijoh, Y., Adachi, H., Sakuma, R., Yeo, C.Y., Yashiro, K., Watanabe, M., Hashiguchi, H., Mochida, K., Ohishi, S. and Kawabata, M. (2000). Left-right asymmetric expression of lefty2 and nodal is induced by a signaling pathway that includes the transcription factor FAST2. Molecular Cell 5, 35-47.
- Cheng, A.M.S., Thisse, B., Thisse, C. and Wright, C.V.E. (2000). The lefty-related factor xatv acts as a feedback inhibitor of nodal signaling in mesoderm induction and l-r axis development in Xenopus. Development 127, 1049-1061.
- Bisgrove, B.W., Essner, J.J. and Yost, H.J. (1999). Regulation of midline development by antagonism of lefty and nodal. Development 126, 3253-3262.
- Capdevila, J., Vogan, K.j., Tabin, C.J. and Izpisua Belmonte, J.C. (2000). Mechanisms of left-right determination in vertebrates. Cell 101, 9-21.