Hierarchical linkage of pattern-forming reactions to obtain a precise number of segments
Based on the pioneering paper of Nüsslein-Volhard and Wieschaus [1] I proposed a model [2,3] for the segmentation in Drosophila (Meinhardt, 1985, 1986). This model was published before the overwhelming amount of molecular-genetic data became available. Nevertheless, the basic stipulations of the model turned out to be correct. Thus, the theory is convenient to illustrate the principles of how to couple several pattern forming systems to get step by step a more complex pattern. Meanwhile also more detailed theoretical studies [5-7] have been published.


A fine-grained pattern by a set of nested pattern forming reaction
Initially, a relatively crude pattern is formed that controls the formation of the next finer pattern, and so on. Imagine that under the influence of the maternal positional information a primary basic subdivision into relative coarse regions is achieved by regional activation of the gap genes. To avoid confusion by the strange and arbitrary names (such as 'krüppel', 'knirps'…) they should be called I, II, III and IV. These regions would be in itself more or less homogeneous. It may appear straightforward to assume that each of these regions become further subdivided into two or three sub-regions. However, the polarity must be transmitted throughout the cascade in order to make sure that the final segments have the correct polarity. Each of the region regions I, II, III... has per se no polarity but the borders between the regions have. For instance, II is anterior to III). For this reason, I have proposed that not the regions itself but the borders between these regions, resulting for instance from an overlap of the signals produced in the regions I and II, II and III... act as a scaffold and organize the subsequent pattern. This has led to the prediction (Meinhardt, 1985) [PDF] that the region in which a gap gene is transcribed is only half as large as the gap in the sequence of segments that is caused by the corresponding mutation. For instance, if the gene responsible for region III is broken, neither the II/III border nor the III/IV border is present. Thus, in addition to the III region, half of the II region and half of the IV region would be lost too. The resulting gap would be twice as large as the III region itself. This prediction has been shown to be correct. The Krüppel gene, for instance, is expressed in a region of about 3.5 future segments [8] while in a Krüppel mutant at least 7 segments are missing.
Pair rule patterns: repetition of at least four elements
As mentioned on the previous page, a periodic structure with polarity requires the repetition of at least three elements. In the pair rule mutants, the remaining pattern still maintains the original polarity. For this reason I have proposed that the pair rule pattern results from the repetition of at least four basic building blocks. If one of the four elements is missing, three elements can remain and the polarity is maintained. Although more than 8 pair rule genes are known, it has turned out that only a mutation in four of them leads to a change in the gene expression of the remaining ones [15], indicating that these genes generate the primary pair rule pattern.
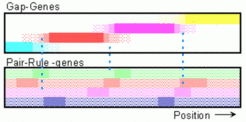
The theory has predicted that the pair rule genes are selectively activated of two gap genes. Now we know that this overlapping signals result from the diffusion of the gap gene proteins. The simulation above shows first the formation of the gap gene pattern. The first pair rule genes become activated around the gap gene border (see snapshot at right). To maintain the polarity, more than one gene has to become activated around each border. (Inspired by the experiments of Klaus Sander with non-Drosophila insects, I assumed the high point of the positional information at the posterior pole).
This prediction that at least two gap gene products are involved in the activation of a pair rule stripe has been experimentally verified [9-11]. A further prediction was that a particular region of gap gene expression on its own is insufficient to give rise to a periodic pattern (since a single region does not contain borders). This is in agreement with the observation. If all maternal positional information is removed by mutations, the gap gene Krüppel is activated everywhere in the egg, but no periodic pattern is formed [12].
On the other hand, from the model I had expected that only few elements of the periodic pattern are fixed by the hierarchically higher gap gene pattern and that the remaining pattern becomes filled in due to the self-organizing capacity of the pair rule pattern. Experiments indicate, however, that the overlapping gap gene pattern is used for a very detailed initiation of the pair rule pattern [13,14]. For the same reason, the gap mutants are not correctly described: Missing segments would form in the course of time, although in a disorganized way.
As expected from the model, at least two of the pair rule genes have a strong feedback on their own activation. However, no long-ranging substances are yet known that could be the signal for the spacing of the stripe-like gene expression. Therefore, it is not yet clear whether there is a direct communication between adjacent pair rule gene activities, analogous to the hedgehog-communication between engrailed and wingless.
For the activation of the segment polarity genes by the pair rule genes, a doubling in the spatial periodicity is required. The same cell state must be activated at two different positions in the double segment pattern. The simulation below shows an animated simulation.


Recent investigations have shown that a stripe-like expression of pair rule genes (hairy) forms also in short germ insects [15]. In these systems the pattern forms after cellularization.
For more details see Meinhardt, (1986) [PDF] ; [5-7] for more recent models. The stripes in Drosophila are certainly not the result of a simple Turing-like instability, since this would never lead to exactly 14 stripes in only a part of the embryo. It was the claim of the experimentalists that the theoreticians were wrong in this respect (see, e.g., [4]).
Further Reading and References
H.M.. (1985). Mechanisms of pattern formation during development of higher organisms: a hierarchical solution of a complex problem. Ber. Bunsenges. Phys. Chem. 89, 691-699 [PDF].
H.M. (1986). Hierarchical inductions of cell states: a model for segmentation in Drosophila J. Cell Sci. Suppl. 4, 357-381 [PDF]
- Nüsslein-Volhard, C. and Wieschaus, E. (1980). Mutations affecting segment number and polarity in Drosophila. Nature 287, 795-801
- Driever, W. and Nüsslein-Volhard, C. (1988). A gradient of bicoid protein in Drosophila embryos. Cell, 54, 83-93.
- Nüsslein-Volhard,C, Frohnhöfer, H.G. and Lehmann R. (1987). Science 238, 1675-1681
- Akam, M. (1989). Drosophila development - making stripes inelegantly. Nature 341, 282-283.
- von Dassow, G., Meir, E., Munro, E.M. and Odell, G.M. (2000). The segment polarity network is a robust development module. Nature 406, 188-192.
- Sanchez, L and Thieffry, D.,(2001). A logical analysis of the Drosophila gap-gene system J. Theor. Biol. 211,115
- Jaeger et al., (2004).Dynamic control of positional information in the early Drosophila embryo. Nature 430, 368-371
- Knipple, D.C., Seifert, E., Rosenberg, U.B., Preiss, A. and Jäckle, H. (1985). Spatial and temporal pattern of Krüppel gene expression in early Drosophila development. Nature 317, 40-44.
- Hülskamp, M. and Tautz, D. (1991). Gap genes and gradients - the logic behind the gaps. BioEssays 13, 261-268.
- Pankratz, E., Seifert, E., Gerwin, N., Billi, B., Nauber, N. and Jäckle, H. (1990). Gradients of Krüppel and knirps gene products direct pair rule gene stripe patterning in the posterior regions of the Drosophila embryo. Cell 61, 309-316.
- Howard, K.R. and Struhl, G. (1990). Decoding positional information - regulation of the pair-rule gene hairy. Development 110, 1223-1231.
- Gaul, U. and Jäckele, H. (1987). Pole region-dependent repression if the Drosophila gap gene Krüppel by maternal gene products. Cell 51, 549-555.
- Carroll, S.B. and Scott, M.P. (1985). Localization of the fushi tarazu protein during Drosophila embryogenesis. Cell 43, 47-57.
- Howard K, Ingham P and Rushlow C (1988). Region-specific alleles of the Drosophila segmentation gene hairy. Genes Dev. 2, 1037-1046.
- Eckert C., Aranda M., Wolff C. and Tautz D. (2004). Separable stripe enhancer elements for the pair-rule gene hairy in the beetle tribolium. EMBO Rep 5,638-642.