Somite formation: sequential conversion of a periodic pattern in time into a periodic pattern in space
Somites are the most obviously segmented structure in vertebrates. Their formation occurs in a different way. Pairs of anterior and posterior half-somites become separated form non-segmented presomitic mesoderm in an anterior-to posterior sequence. The drawings show a chicken embryo at 25h and 37h with 5 or 12 somites respectively:

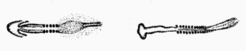
To be compatible with classical observations, the counter-intuitive prediction was made that the specification of anterior and posterior half-somites occur by wave-like processes that are initiated at the posterior end of the presomitic mesoderm, move towards anterior and sharpens until they comes to rest at a given distance from the last formed half-somite (Meinhardt, 1982 , 1986b). A description of the model including animated simulations is also available in a PowerPoint presentation [PPT]. The prediction has been confirmed by the observation of the c-hairy1 gene in the chicken (Palmeirim et al., 1997), [2].
While, as explained above, anterior (A) and posterior (P) cells stabilize each other in the region of a common boundary, it is a property of such an interaction that a group of cells consisting of one type only (A or P) can oscillate back and forth between the two states. For instance, if all cells are in A-state, the P-state obtains substantial help while the A-state is not supported. Therefore, after a certain time, the cells switch from A to P. Later, for the same reason, the cells switch back to A, and so on.

Such a spatially homogeneous oscillating system can convert into a periodic pattern that is stable in time if, at any location, an A- P border has been formed. Imagine that all cells are in the P state except of the most anterior cells that are in the A state. Cells close to this border stabilize each other while the P cells distant to this border switch from P to A, forming in this way a second A/ P border, and so on. As shown in the simulation above, the periodic pattern can be elongated by terminal growth.
But what gives rise to the first border? To account for several observations, it is assumed that some sort of positional information provides the driving force for this oscillation. Imagine a field of cells exposed to a graded morphogen concentration, which has its high point at the posterior end, and further, that all cells are in the P state. Cells above a certain threshold concentration switch from P to A, a process that leads to the first P- A border. Again, cells distant to the new P/ A border will switch back to A, forming the next A/ P border, and so on. After each full cycle, one pair of A/P stripes is added. In the course of time, the region of the stable spatially alternating A- P pattern enlarges at the expense of the cells that still oscillate between A and P. The borderline between the stable and oscillating cells move over the field in a wave-like manner. Since the oscillation is driven by a gradient with a high point at the posterior end, the transitions occurs a bit earlier there and sweeps towards anterior in a wave-like manner. It comes to rest at a certain distance from the last-formed border. Thus, although the somites separate from each other in an anterior to posterior sequence, it was predicted that this is based on oscillations with a wave-like phase shift causing an apparent wave in posterior-to anterior direction. The dynamics of the c-hairy1 expression pattern as observed by Palmeirim et al. [2] in chickens corresponds the expected pattern for the P-state.

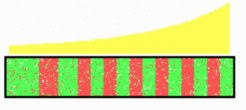
In addition to the generation of the periodic pattern, the oscillations are proposed to have another function. As can be seen from the simulation above, each more posterior somite was going through exactly one more A/P cycle. This allows a sort of digital counting on the gene level: each full cycle of this oscillation advances the specification of the presomitic mesoderm towards being more posterior. Also this prediction has found recently strong support: the activation of the HOX genes is under control of the segmental clock. [3-5]

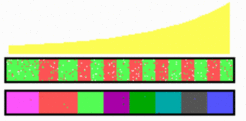
In the simulation above, each transition from anterior to posterior half somite specification (green-red transition) enables the activation of a subsequent gene (specifying a more posterior structure). In the course of time, each pair of A/P half somites one particular gene is active.
Meanwhile it has turned out that not every pair of half-somites is characterized by a particular HOX gene activation. The number of somites is highly variable. In terms of the model, it may require several full cycles before the next HOX gene becomes activated. The idea is that some sort of isolators acts as stop points on the activation of a subsequent HOX gene. Each full cycle overcomes one of these stop points. The number of periodic structures (somites) in which a particular HOX gene is active depends on the number of such stop points. They can be easily changed by mutations.
Such a coupling between a periodic pattern and a sequential pattern formation is also required for the short germ insects mentioned above, since the transition from one compartmental specification to the next and the activation of new HOX genes occurs precisely in register. Since mutations in the HOX genes change only the quality of the segment, not the segmentation as such, the generation of the periodic pattern is obviously the primary event. This makes also sense in evolutionary terms according to which the formation of identical units was a primary step. These units became different from each other in a second step. In the following simulation an alteration between three cell states take place (upper lane). After a full cycle, a subsequent specifying (HOX-) gene becomes activated. Both the sequential and the periodic pattern are precisely in register.


Further Reading and References
The conversion of a periodic pattern in time into a periodic pattern in space was first published in the book (Meinhardt, 1982) "Models of biological pattern formation", chapter 14 [PDF] . A slighly modified version appered in a proceeding volume devoted to somite formation [PDF] (Models of segmentation. In: Somites in developing embryos (R.Bellairs, D.A.Edie, J.W. Lash, Edts), Nato ASI Series A, Vol 118, pp 179-189, Plenum Press, New York).
- Palmeirim, I., Henrique, D., Ishhorowicz, D. and Pourquie, O. (1997). Avian hairy gene-expression identifies a molecular clock linked to vertebrate segmentation and somitogenesis. Cell 91, 639-648.
- Dale, K. and Pourquie, O. (2000). A clock-work somite. BioEssays 22, 72-83.
- Zakany, J., Kmita, M., Alarcon, P., de la Pompa, J.L. and Duboule, D. (2001). Localized and transient transcription of hox genes suggests a link between patterning and the segmentation clock. Cell 106, 207-217.
- Dubrulle, J., McGrew, M.J. and Pourquie, O. (2001). Fgf signaling controls somite boundary position and regulates segmentation clock control of spatiotemporal hox gene activation. Cell 106, 219-232.