The dorsal organizer and its A-P polarity
Crucial for the dorsoventral patterning is the BMP / Chordi n interaction (Dpp / short gastrulation in Drosophila). For some systems there are indications components have properties as we expect for pattern-forming systems. BMP and Chordin inhibit each other ([8-12], for review (13,14]). Two components that mutually inhibit each other behave as if positive autoregulation is involved. For instance, an increase of the first component leads to an enforced repression of the second, which, in turn, leads to a further increase of the first as if this substance would be directly positively autoregulating. To obtain a balanced activation of BMP and Chordin at opposite positions, a third component is anticipated that acts antagonistically on one of these indirectly self-enhancing reactions. A candidate is the Anti-Dorsalizing Morphogenetic Protein (ADMP) [15]. Its properties have been frequently regarded as counterintuitive: being expressed in the organizer, but its function is to reduce organizer activity. However, as its acts over a longer range [16,17], it satisfies the theoretical expectations: being produced in the organizer region and antagonizing a self-enhancing reaction. The simulation below shows a pattern formation based on an indirect autocatalysis of Chordin (green) and BMP (blue). ADMP (red) is produced only in the green area but distributes over a larger area. Since Chordin (green) is producing the antagonist, the Chordin region is more confined. In this simulation, no maternally supplied asymmetries are assumed. Therefore, it takes some times until all components have obtained their final localization.

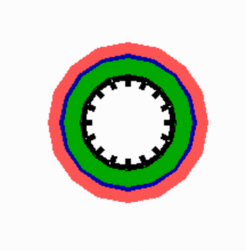
Alternatively, BMP may be in itself a component of a pattern-forming system since the condition for autoregulation is satisfied (Hild et al., 1999). Also a secreted BMP antagonist has been found that is produced under BMP control; Sizzled [18]. Thus, both the organizer and the antipodal side may be under control of two genuine pattern-forming reactions that keep distance by mutual exclusion. Such a scheme accounts for the fact that a residual pattern formation takes place even if one or the other system is non-functional.
The Spemann organizer is from early stages onwards subdivided into head and trunk organizer, with goosecoid (gsc) being a marker for the head organizer. This gene can be activated by high concentrations of TGF-ß factors such as Activin, Xnr1, Xnr2 and Xnr4. A threshold concentration is required for its activation, but once activated it remains so independent of the signal. Based on these facts a modified version of the "three-factor model" (see [1]) was proposed [2], in which the (eccentric) Nieuwkoop center produces higher amounts of TGF-ß like factors that diffuse to the mesoderm. Therefore, a gradient generated by a source region in the endoderm induces gene activations in the mesoderm in a concentration-dependent manner with gsc at its highest concentrations within the mesoderm. In the chick, a similar early subdivision of the organizer, Hensens's node, into adjacent anterior and posterior portions has been observed [3]. It takes place under the influence of the posterior marginal zone, the equivalent of the Nieuwkoop center.

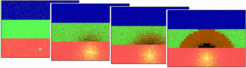
[Movie]
A simulation of the formation the Spemann organizer with its A-P polarity’s given above. The simulation shows a dorsal view of the equatorial zone. Assumed is a preceding ecto/meso/endo subdivision (red/green, blue, see simulation at the preceding page). An organizing region displaced from the vegetal pole is formed in the endoderm and marks the future dorsal side, the Nieuwkoop center. A long ranging signal (yellow) that spreads from this center causes a promotion of mesodermal cells: first an activation of genes for trunk organizer occurs (brown; e.g., flh). In regions in which the concentration is sufficiently high, genes for head formation (black, e.g. goosecoid) become activated, a process that is connected with the suppression of the trunk-specific genes. Thus, genes such as goosecoid become activated closer to the endoderm. Since these cells involute first, they become localized most closely to the animal pole, forming the most anterior part of the organism.
The model for the step-wise activation of genes under the influence of a morphogen signal (see Gene Activation ) predicted particular regulatory features. After promotion is completed, a subsequent increase of the signal can lead to a further promotion In contrast, a decrease of the signal is without effect because each transition from one gene activation to the next is essentially irreversible ("distal transformation", step-wise promotion). The activation of brachyury and goosecoid by different concentrations of Activin [4] shows this behavior. Gurdon et al. [4] came to the same conclusion, i.e., that step-wise promotion takes place and called this a “ ratchet-like" transition. In the zebra fish, low concentrations of the nodal-related factors cyclops and squint activate floating head, a gene that is required for notochord formation in the trunk. Higher concentrations leads to goosecoid activation [5]. If goosecoid activation is abolished by blocking a necessary co-factor, the expression of the gene that is activated already at low concentrations expands into the region in which normally the "high" gene is expressed, i.e., notochord enlarges on the expense of prechordal plate, as expected. From the model it is expected that each of these genes have positive and negative autoregulatory elements. Indications for both have been found for goosecoid [6,7].
Further Reading and References
- Heasman, J. (1997). Patterning the Xenopus blastula. Development, 124, 4179-4191.
- Agius, E., Oelgeschlager, M., Wessely, O., Kemp, C. and De, R.E. (2000). Endodermal nodal-related signals and mesoderm induction in Xenopus. Development 127, 1173-1183
- Bachvarova, R.F., Skromne, I. and Stern, C.D. (1998). Induction of primitive streak and Hensen's node by the posterior marginal zone in the early chick embryo. Development 125, 3521-3534.
- Gurdon, J.B., Harger, P., Mitchell, A. and Lemaire, P. (1994). Activin signalling and response to a morphogen gradient. Nature 371, 487-492.
- Gritsman, K., Talbot, W.S. and Schier, A.F. (2000). Nodal signaling patterns the organizer. Development 127, 921-932.
- Blumberg,B,Wright,CVE,De_Robertis,EM,Cho,KWY (1991). Organizer-specific homeobox genes in Xenopus laevis embryos. Science 253,194-196
- Danilov, V., Blum, M., Schweickert, A., Campione, M. and Steinbeisser, H. (1998). Negative autoregulation of the organizer-specific homeobox gene goosecoid. J. Biol. Chem. 273, 627-635.
- Piccolo, S., Agius, E., Lu, B., Goodman, S., Dale, L. and De Robertis, E.M. (1997). Cleavage of Chordin by Xolloid metalloprotease suggests a role for proteolytic processing in the regulation of Spemann organizer activity. Cell 91, 407-416.
- Blitz, I.L., Shimmi, O., Wunnenberg-Stapleton, K., O'Connor, M.B. and Cho, K.W.Y. (2000). Is Chordin a long-range- or short-range-acting factor? roles for BMP1-related metalloproteases in Chordin and BMP4 autofeedback loop regulation. Dev. Biol. 223, 120-138.
- Jones, C. M. and Smith, J.C. (1998). Establishment of a BMP-4 morphogen gradient by long-range inhibition. Dev. Biol. 194, 12-17
- Balemans, W. and Van Hul, H.W. (2002). Extracellular regulation of BMP signaling in vertebrates: a cocktail of modulators. Dev. Biol 250, 231-250.
- Sasai, Y. (2001). Regulation of neural determination by evolutionarily conserved signals: anti-bmp factors and what next?. Current Opinion in Neurobiology 11, 22-26.
- Harland, R. and Gerhart, J. (1997). Formation and function of Spemann's organizer. Ann. Rev. Cell Dev. Biol. 13, 611-667.
- De Robertis, E.M., Larraín, J, Oelschläger, M. and Wessely, O. (2000). The establishment of Spemann's organizer and patterning of the vertebrate embryo. Nature Rev. Genetics 1, 171-181.
- Moos, M., Wang, S.W. and Krinks, M. (1995). Anti-dorsalizing morphogenetic protein is a novel tgf-beta homolog expressed in the Spemann organizer. Development 121, 4293-4301.
- Lele, Z., Nowak, M. and Hammerschmidt, M. (2001). Zebrafish ADMP is required to restrict the size of the organizer and to promote posterior and ventral development. Dev. Dyn. 222, 681-687
- Willot, V., Mathieu, J., Lu, Y., Schmid, B., Sidi, S., Yan, Y.L., Postlethwait, J.H., Mullins, M., Rosa, F. and Peyrieras, N. (2002). Cooperative action of ADMP- and BMP-mediated pathways in regulating cell fates in the zebrafish gastrula. Dev. Biol. 241, 59-78.
- Martyn, U. and Schulte-Merker, S. (2003). The ventralizing ogon mutant phenotype is caused by the zebrafisch homologue of Sizzled, a secreted Frizzled-related protein. Dev. Biol. 260, 58-67