The transition from the radial- to the bilateral-symmetric body plan:
An ancestral body pattern evolved into brain and heart of higher organisms
Hydra provides a key for the assignment of ancestral and contemporary body axes
Simple radial symmetric animals have only a single axis (oral-aboral) and a single opening of the gastric cavity, the blastopore. A central issue of evolutionary biology is how the two major body axes of higher organisms, anteroposterior and dorsoventral, evolved from such an ancestral body plan. In the patterning of the freshwater polyp hydra, molecules have been found that participate in higher organisms in the generation of both axes. Thus, hydra and other Coelenterates are assumed to provide information about the relation of the ancestral single axis and the two main axes of bilateral-symmetric organisms.
Problems with the conventional protostomia-deuterostomia concept
Two conflicting hypothesis have been put forward. According to the protostomia - deuterostomia concept, a widening and deformation of the blastopore took place in one dimension, resulting in a slit-like geometry that forms the future midline (below left). In protostomia such as insects, the anterior part of the slit remained open as the mouth. In deuterostomia including vertebrates, a posterior part remained open forming the anus.


The alternative hypothesis is that the blastopore represents the most posterior part of an organism (figure top right). This view is strongly supported be recently observed expression patterns in hydra. This implicates, however, that a separate signalling system was required that allow the generation of an organizing region for the dorsoventral patterning. The generation of such a midline organizer with a long extension in one (anteroposterior) and a short extension perpendicular to it is an intricate problem.
The system that was patterning once the bodies of radial-symmetric ancestors was later used to pattern essentially the head and the heart of higher organisms
As indicated by the expression pattern of Wnt - and Brachyury, the gastric opening in hydra, (hypostome) represents the most posterior part of the organisms although this part, the hypostome with the surrounding tentacles, is conventionally called the ‘head’. If this assigmment is correct, the antipodal (aboral) part, the so-called foot is homologous to the most anterior structures: Indeed, Nkx2.5 is expressed in the foot of hydra [3] and in the forebrain and heart of higher organisms (pink in the figure below). The tentacle/hypostome border has the same positional address as the midbrain/hindbrain border in vertebrates (posterior border of Otx-expression [2]). The recently observed highly conserved patterning in the brain of higher organisms is proposed to have its origin in the preserved body pattern of radial-symmetric common ancestors (for details and references see Meinhardt, 2002).

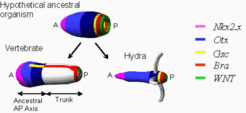
The anteroposterior axis consists of two parts; the trunk is evolutionary a later invention
With the specialization of the anterior portion towards brain function, the trunk, the largest portion of higher animals, was added during further evolution, for instance, to enable an efficient locomotion. The corresponding zone where this took place can be precisely localized in hydra. In a narrow zone between the tentacles and the gastric opening Goosecoid (yellow) [6] and the Brachyury (red) [4] homologues are expressed in adjacent regions. Since in vertebrates Goosecoid is involved in head patterning, Brachyury in tail formation, the trunk evolved and differentiated next to the posterior end of the ancestral organism. This sequence is recapitulated in the ontogenetic development of many animals including vertebrates, short germ insects and annelids in which the head is formed first and the trunk is added subsequently. In hydra Goosecoid and Brachyury are not yet separated by the trunk. This agrees with the observation that the typical HOX genes characteristic for the trunk in higher organisms with their 3'-5' co-linear arrangement have not been found in hydra. This supports the view that hydra preserves an ancestral axial pattern [7] before trunk formation and bilaterality evolved.
The foot can be regarded as an ancestral heart. The heart of higher organisms and the foot of hydra are under the control of the same gene, NKx2.5. Both are initiated at the same (very anterior) position, have pumping activity, and the constriction frequency of both can be influenced by the same peptide [1].
Further Reading and References
Meinhardt,H (2002). The radial-symmetric hydra and the evolution of the bilateral body plan: an old body became a young brain. BioEssays 24, 185-191. [PDF]
- Shimizu H, Fujisawa T: Peduncle of Hydra and the heart of higher organisms share a common ancestral origin. Genesis 2003
- Smith, K. M., Gee, L., Blitz, I. L. and Bode, H.R. (1999). CnOtx, a member of the Otx gene family, has a role in cell movement in hydra. Dev. Biol. 212, 392- 404.
- Grens, A., Gee, L., Fisher, D.A. & Bode, H.R. (1996). Cnnk-2, a nk-2 homeobox gene, has a role in patterning the basal end of the axis in hydra. Dev. Biol. 180, 473-488.
- Technau, U. & Bode, H.R. (1999). HyBra1, a Brachyury homologue, acts during head formation in Hydra. Development 126, 999-1010.
- Hobmayer etal., (2000) Wnt signalling molecules act in axis formation in the diploblastic metazoan hydra. Nature 407, 186-189.
- Broun, M., Sokol, S. & Bode, H.R. (1999). Cngsc, a homologue of goosecoid, participates in the patterning of the head, and is expressed in the organizer region of hydra. Development 126, 5245-5254.
- Ball, E.E., Hayward, D.C., Saint,R. and Miller,D.J. (2004). A simple plan: cnidarians and the origins of developmental mechanisms. Nature Rev, Genetics 5,567 -577